
Восстановление
рис. 19, А). Главным механизмом утомления в этих упражнениях служат связанное с интенсивным гликогено-лизом (как основным, путем энергопродукции) накопление лактата в мышцах (см. рис. 19, Б) и крови и обусловленное им снижение рН в мышечных клетках и крови.
Оба эти фактора приводят к уменьшению скорости гликогенолиза в мышцах и оказывают отрицательное влияние на деятельность ЦНС. При работе субмаксимальной анаэробной мощности дополнительным (хотя не очень существенным) фактором, лимитирующим работоспособность, служат функциональные возможности, кислородтранспортной системы.
Поэтому одним из механизмов утомления при выполнении такой работы является недостаточное снабжение мышц кислородом.
При выполнении упражнений максимальной аэробной мощности утомление связано прежде всего с кислородтранспортной системой, предельные возможности которой являются фактором, лимитирующим работоспособность. Один из главных механизмов утомления в данном случае - недостаточное обеспечение работающих: мышц кислородом.
В процессе такой работы значительную долю энергии-мышцы получают в результате анаэробного гликогенолиза с образованием молочной кислоты, накопление которой (снижение рН) в мышцах и крови также играет важную роль в развитии утомления.
Выполнение упражнений околомаксимальной аэробной мощности также лимитируется в основном возможностями кислородтранспортной системы. В процессе их выполнения концентрация фосфа-генов снижается незначительно, концентрация лактата в мышцах и крови относительно невелика (см. рис. 19).
Утомление связано со снижением производительности сердечно-сосудистой системы, особенно сердца. Сердечная производительность выступает как главный фактор, лимитирующий снабжение мышц кислородом. Работа обеспечивается преимущественно гликогенолизом.
Однако отказ от продолжения ее прямо не связан с истощением углеводных ресурсов организма (см. рис. 20). Высокая концентрация молочной кислоты в мышцах и крови позволяет рассматривать ее как один из важных механизмов утомления при выполнении упражнений околомаксимальной аэробной мощности,
Упражнения субмаксимальной аэробной мощности связаны с большой нагрузкой на сердечнососудистую систему. Их выполнение-обеспечивается окислительными процессами в работающих мыцн цах, использующих в качестве основного субстрата мышечный гликоген и глюкозу крови. Главным механизмом утомления при таких упражнениях служит истощение запасов гликогена в работающих мышцах и печени (см. рис. 20).
Большинство изменений, наблюдаемых в деятельности сердечно-сосудистой системы, на протяжении периода квазиустойчивого состояния (см. рис. 13), отражает течение процессов, которые в конце концов приводят к утомлению. Большая и длительная нагрузка на сердце ведет к снижению производительности миокарда.
Определенную роль в развитии утомления играют повышающиеся по мере продолжения работь! требования к поддержанию необходимой температуры тела (рабочей гипертермии).
Упражнения средней аэробной мощности также оказывают наибольшую нагрузку на кислородтранспортную систему. При работе такой мощности происходит значительный расход гликогена мышц и усиленный расход (истощение) гликогена печени, что ведет к развитию гипогликемии. Таким образом, вторично страдает ЦНС, для которой глюкоза крови играет роль единственного энергетического источника.
Кроме того, большое значение имеет нарушение процессов терморегуляции, что может вызвать критическое повышение температуры тела. В результате дополнительного перераспределения кровотока (усиления кожного кровотока и снижения кровотока работающих мышц) происходит повышение теплоотдачи.
Доставка кислорода к рабочим мышцам снижается, что ведет к мышечному утомлению.
Упражнение малой аэробной мощности в значительной Мере характеризуются теми же локализацией и механизмами утомления, что и упражнения средней аэробной мощности. Отличие состоит в более медленном наступлении описанных процессов и в большем расходовании жиров, недоокисленные продукты расщепления которых могут поступать в кровь и быть важным фактором утомления.
Восстановление
После прекращения упражнения происходят обратные изменения в деятельности тех функциональных систем, которые обеспечивали выполнение данного упражнения. Вся совокупность изменений в этот период объединяется понятием восстановление. На протяжении восстановительного периода удаляются продукты рабочего метаболизма и восполняются энергетические запасы, пластические (структурные) вещества (белки и др.) и ферменты, израсходованные за время мышечной деятельности.
По существу, происходит восстановление нарушенного работой гомеостазга. Однако восстановление - это не только процесс возвращения организма к предрабочему состоянию.
В этот период происходят также изменения, которые обеспечивают повышение функциональных возможностей организма, т. е. положительный тренировочный эффект.
Восстановление функций после прекращения работы
Сразу после прекращения работы происходят многообразные изменения в деятельности' различных функциональных систем. В периоде восстановления можно выделить 4 фазы: 1) быстрого восстановления, 2) замедленного восстановления, 3) суперкомпенсации (или перевосстановления), 4) длительного (позднего) восстановления.
Наличие этих фаз, их длительность и характер сильно варьируют для разных функций. Первым двум фазам соответствует период восстановления работоспособности, сниженной в результате утомительной работы, третьей фазе - повышенная работоспособность, четвертой - возвращение к нормальному (предрабочему) уровню работоспособности (рис.
23).
Общие закономерности восстановления функций после работы состоят в следующем. Во-первых, скорость и длительность восстановления большинства функциональных показателей находятся в прямой зависимости от мощности работы: чем выше мощность работы, тем большие изменения происходят за время работы и (соответственно) тем выше скорость восстановления. Это означает, что чем короче предельная продолжительность упражнения, тем короче период восстановления.
Так, продолжительность восстановления большинства функций после максимальной анаэробной работы - несколько минут, а после продолжительной работы, например после марафонского бега, - несколько дней. Ход начального восстановления многих функциональных показателей по своему характеру является зеркальным отражением их изменений в период
Во-вторых, восстановление различных функций протекает с разной скоростью, а в некоторые фазы восстановительного процесса и с разной направленностью, так что достижение ими уровня покоя происходит неодновременно (гетерохронно). Поэтому о завершении процесса восстановления в целом следует судить не по какому-нибудь одному и даже не по нескольким ограниченным показателям, а лишь по возвращению к исходному (предрабочему) уровню наиболее медленно восстанавливающегося показателя (М.
Я. Горкин).
В-третьих, работоспособность и многие определяющие ее функции организма на протяжении периода восстановления после интенсивной работы не только достигают предрабочего уровня, но могут и превышать его, проходя через фазу перевосстановления (см. рис. 23). Когда речь идет об энергетических субстратах, то такое временное превышение предрабочего уровня носит название суперкомпенсации (Н.
Н. Яковлев).
Кислородный долг и восстановление энергетических запасов организма
В процессе мышечной работы расходуются кислородный запас организма, фосфагены (АТФ и КрФ), углеводы, (гликоген мышц и печени, глюкоза крови) и жиры. После работы происходит их восстановление.
Исключение составляют жиры, восстановления которых может и не быть.
Восстановительные процессы, происходящие в организме после работы, находят свое энергетическое отражение в повышенном (п сравнению с предрабочим состоянием) потреблении кислорода - кислородном долге (см. рис. 12). Согласно оригинальной теории А. Хйлла (1922), кислородный долг - это избыточное потребление О2 сверх предрабочего уровня покоя, которое обеспечивает энергией организм для восстановления до предрабочего состояния, включая восстановление израсходованных во время работы запасов энергии и устранение молочной кислоты.
Скорость потребления О2 после работы снижается экспоненциально: на протяжении первых 2-3 мин очень быстро (быстрый, или алактатньш, компонент кислородного долга), а затем более медленно (медленный, или лактатный, компонент кислородного долга), пока не достигает (через 30-60 мин) постоянной величины, близкой к предрабочей.
После работы мощностью до 60% от МПК кислородный долг не намного превышает кислородный дефицит. После более интенсивных упражнений кислородный долг значительно превышает кислородный дефицит, причем тем больше, чем выше мощность работы (рис.
24).
Быстрый (алактатный) компонент О2-долга связан главным образом с использованием О2 на быстрое восстановление израсходованных за время работы высокоэнергетических фосфагенов в рабочих мышцах, а также с восстановлением нормального содержания О2 в венозной крови и с насыщением миоглобина кислородом.

Медленный (лактатный) компонент О2-долга связан со многими факторами. В большой мере он связан с после-рабочим устранением лактата из крови и тканевых жидкостей.
Кислород в этом случае используется в окислительных реакциях, обеспечивающих ресинтез гликогена из лактата крови (главным образом, в печени и отчасти в почках) и окисление лактата в сердечной и скелетных мышцах. Кроме того, длительное повышение потребления О2 связано с необходимостью поддерживать усиленную деятельность дыхательной и сердечно-сосудистой систем в период восстановления, усиленный обмен веществ и другие процессы, которые обусловлены длительно сохраняющейся повышенной активностью симпатической нервной и гормональной систем, повышенной температурой тела, также медленно снижающимися на протяжении периода восстановления.
Восстановление запасов кислорода. Кислород находится в мышцах в форме химической связи с миоглобином.
Эти запасы очень невелики: каждый килограмм мышечной массы содержит около 11 мл О2. Следовательно, общие запасы мышечного кислорода (из расчета на 40 кг мышечной массы у спортсменов) не превышают 0,5 л. В процессе мышечной работы он может быстро расходоваться, а после работы быстро восстанавливаться.
Скорость восстановления запасов кислорода зависит лишь от доставки его к мышцам.
Сразу после прекращения работы артериальная кровь, проходящая через мышцы, имеет высокое парциальное напряжение (содержание) О2, так что восстановление О2-миоглобина происходит, вероятно, за несколько секунд. Расходуемый при этом кислород составляет некоторую часть быстрой фракции кислородного долга, в которую входит также небольшой объем О2 (до 0,2 л), идущий, на восполнение нормального содержания его в венозной крови.
Таким образом, уже через несколько секунд после прекращения работы кислородные запасы в мышцах и крови восстанавливаются. Парциальное напряжение О2 в альвеолярном воздухе и в артериальной крови не только достигает предрабочего уровня, но и превышает его. Быстро восстанавливается также содержание О2 в венозной крови, оттекающей от работавших мышц и других активных .органов и тка-'ней тела, что указывает на достаточное их обеспечение кислородом в послерабочий период.
Поэтому нет никаких физиологических оснований использовать дыхание чистым кислородом или смесью с повышенным содержанием кислорода после работы для ускорения процессов восстановления.
Восстановление фосфагенов (АТФ и КрФ). Фосфагены, особенно АТФ, восстанавливаются очень быстро (рис. 25). Уже на протяжении 30 с после прекращения работы восстанавливается до 70% израсходованных фосфагенов, а их полное восполнение заканчивается за несколько минут, причем почти исключительно за счет энергии аэробного метаболизма, т. е. благодаря кислороду, потребляемому в быструю фазу О2-долга.
Действительно, если сразу после работы жгутировать работающую конечность и таким образом лишить мышцы кислорода, доставляемого с кровью, то восстановление КрФ не произойдет.
Чем больше расход фосфагенов за. время работы, тем больше требуется О2 для их восстановления (для восстановления 1 моля АТФ необходимо 3,45 л О2). Величина быстрой (алактатной) фракции О2-долга прямо связана со степенью снижения фосфагенов в мышцах к концу работы.
Поэтому данная величина указывает на количество израсходованных в процессе работы фосфагенов.
У нетренированных мужчин максимальная величина быстрой фракции О2-долга достигает 2-3 л. Особенно большие величины этого показателя зарегистрированы у представителей скоростно-силовых видов спорта (до 7 л у высококвалифицированных спортсменов). В этих видах спорта содержание фосфагенов и скорость их расходования в мышцах прямо определяют максимальную и поддерживаемую (дистанционную) мощность упражнения.
Восстановление гликогена. По первоначальным представлениям Р. Маргария и др. (1933), израсходованный за время работы гликоген ресинтезируется из молочной кислоты на протяжении 1-2 ч после работы. Расходуемый в этот период восстановления кислород определяет вторую, медленную, или
лактатную, фракцию О2-Долга. Однако в настоящее время установлено, что восстановление гликогена в мышцах может длиться до 2-3 дней
Скорость восстановления гликогена и количество его восстанавливаемых запасов в мышцах и печени зависит от двух основных факторов: степени расходования гликогена в процессе работы и характера пищевого рациона в период восстановления. После очень значительного (более 3/4 исходного содержания), вплоть до полного, истощения гликогена в рабочих мышцах его восстановление в первые часы при обычном питании идет очень медленно, и для достижения предрабочего уровня требуется до 2 суток.
При пищевом рационе с высоким содержанием углеводов (более 70% суточного калоража) этот процесс ускоряется - уже за первые 10 ч в рабочих мышцах восстанавливается более половины гликогена, к концу суток происходит его полное восстановление, а в печени содержание гликогена значительно превышает обычное. В дальнейшем количество гликогена в рабочих мышцах и в печени продолжает увеличиваться и через 2-3 суток после истощающей нагрузки может превышать предрабочее в 1,5-3 раза - феномен суперкомпенсации (см. рис.
21, кривая 2).
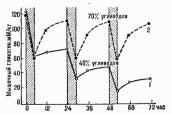
Рис. 26.
Динамика изменения содержания гликогена в рабочих мышцах при ежедневных тренировках (пробегание 16,2 км за час обозначено штриховкой): 1 - с обычным питанием (40% суточного калоража за счет углеводов) и 2 - с повышенным углеводным литанией (70% калоража за счет углеводов) (Д. Костилл, 1976).
При ежедневных интенсивных и длительных тренировочных занятиях содержание гликогена в рабочих мышцах и печени существенно снижается ото дня ко дню, так как при обычном пищевом рационе даже суточного перерыва между тренировками недостаточно для полного восстановления гликогена. Увеличение содержания углеводов в пищевом рационе спортсмена может обеспечить полное восстановление углеводных ресурсов организма к следующему тренировочному занятию (рис.
26).
Устранение молочной кислоты. В период восстановления происходит устранение молочной кислоты из рабочих мышц, крови и тканевой жидкости, причем тем быстрее, чем меньше образовалось молочной кислоты во время работы. Важную роль играет также послерабочий режим.
Так, после максимальной нагрузки для полного устранения накопившейся молочной кислоты требуется 60-90 мин в условиях полного покоя - сидя или лежа (пассивное восстановление). Однако, если после такой нагрузки выполняется легкая работа (активное восстановление), то устранение молочной Кислоты происходит значительно быстрее. У нетренированных людей оптимальная интенсивность восстанавливающей нагрузки - примерно 30-45% от МПК (например, бег трусцой), а у хорошо тренированных спортсменов - 50-60% от МПК, общей продолжительностью примерно 20 мин (рис.
27).

Существует четыре основных пути устранения молочной кислоты: 1) окисление до СО2 и ШО (так устраняется примерно 70% всей накопленной молочной кислоты); 2) превращение в гликоген (в мышцах и печени) и в глюкозу (в печени) -около 20%; 3) превращение в белки (менее 10%); 4) удаление с мочой и потом (12%). При активном восстановлении доля молочной кислоты, устраняемой аэробным путем, увеличивается.
Хотя окисление молочной кислоты может происходить в самых разных органах и тканях (скелетных мышцах, мышце сердца, печени, почках и др.), наибольшая ее часть окисляется в скелетных мышцах ( особенно их медленных волокнах) . Это делает понятным, почему легкая работа (в ней участвуют в основном медленные мышечные волокна) способствует более быстрому устранению лактата после тяжелых нагрузок.
Значительная часть медленной (лактатной) фракции О2-долга связана с устранением молочной кислоты. Чем интенсивнее нагрузка, тем больше эта фракция. У нетренированных людей она достигает максимально 5-10 л, у спортсменов, особенно у представителей скоростно-силовых видов спорта, - 15-20 л.
Длительность ее - около часа. активном восстановлении.
Величина и продолжительность лактатной фракции О2-долга уменьшаются при
Активный отдых
Характер и длительность восстановительных процессов могут изменяться в зависимости от режима деятельности спортсменов в послерабочий, восстановительный, период. В опытах И. М. Сеченова было показано, что в определенных условиях более быстрое и более значительное восстановление работоспособности обеспечивается не пассивным отдыхом, а переключением на другой вид деятельности, т. е. активным отдыхом.
В частности, он обнаружил, что работоспособность руки, утомленной работой на ручном эргографе, восстанавливалась быстрее и полнее, когда период отдыха ее был заполнен работой другой руки. Анализируя этот феномен, И. М. Сеченов предположил, что афферентные импульсы,
поступающие во время отдыха от других работающих мышц, способствуют лучшему восстановлению работоспособности нервных центров, как бы заряжая их энергией. Кроме того, работа одной рукой вызывает увеличение кровотока в сосудах другой руки, что также может способствовать более быстрому восстановлению работоспособности утомленных мышц.
Положительный эффект активного отдыха проявляется не только при переключении на работу других мышечных групп, но и при выполнении той же работы, но с меньшей интенсивностью. Например, переход от бега с большой скоростью к бегу трусцой также оказывается эффективным для более быстрого восстановления. Молочная кислота устраняется из крови быстрее при активном отдыхе, т. е. в условиях работы сниженной мощности, чем при пассивном отдыхе (см. рис.
27). С физиологической точки зрения, положительный эффект заключительной работы невысокой мощности в конце тренировки или после соревнования является проявлением феномена активного отдыха.
Глава 3. Физиологические основы мышечной силы и скоростно-силовых качеств (мощности)
Как уже отмечалось, проявляемая мышечная сила находится в обратной зависимости от скорости движения (см. рис. 1): чем выше скорость движения, тем меньше проявляемая сила, и наоборот.
Разные спортивные упражнения относятся к разным точкам кривой сила - скорость. Упражнения с внешней нагрузкой, близкой или равной максимальной изометрической мышечной силе, относятся к собственно-лиловым упражнениям.
Таковы, например, гимнастические упражнения стойка на кистях, крест, переднее равновесие на кольцах, тяжелоатлетические упражнения со штангой околомаксимального или максимального веса.
При уменьшении внешнего сопротивления скорость движения возрастает, а проявляемая мышечная сила падает. Упражнения с внешней нагрузкой, равной 40-70% от максимальной изометрической силы, при выполнении которых проявляются относительно большие сила и скорость мышечных сокращений, т. е. большая мощность, относятся к скоростно-силовым упражнениям.
Таковы, например, бег на короткие дистанции, прыжки.
В движениях с перемещением малой массы (менее 40% от максимальной изометрической силы) достигается высокая скорость, а проявляемая мышечная сила относительно мала. Такие упражнения относятся к скоростным (например, метание малого мяча с места), движения ненагруженных конечностей).
Границы, разделяющие названные виды упражнений, очень условны.
Физиологические основы мышечной силы
В условиях изометрического сокращения мышцы проявляют максимальную статическую силу.
Максимальная статическая сила и максимальная произвольная статическая сила мышц
Изометрически сокращающаяся мышца развивает максимально возможное для нее напряжение при одновременном выполнении следующих трех условий:
1. активации всех двигательных единиц (мышечных волокон) данной мышцы;
2. режиме полного тетануса у всех ее двигательных единиц;
3. сокращении мышцы при длине покоя.
В этом случае изометрическое напряжение мышцы соответствует ее максимальной статической силе.
Максимальная сила (МС), развиваемая мышцей, зависит от числа мышечных волокон, составляющих данную мышцу, и от их толщины. Число и толщина волокон определяют толщину мышцы в целом, или, иначе, площадь поперечного сечения мышцы (анатомический поперечник). Отношение МС мышцы к ее анатомическому поперечнику называется относительной силой мышцы.
Она измеряется в ньютонах или килограммах силы на 1 см2 (Н/см2 или кг/см2).
Анатомический поперечник определяется как площадь поперечного разреза мышцы, проведенного перпендикулярно к ее длине. Поперечный разрез мышцы, проведенный перпендикулярно к ходу ее волокон, позволяет получить физиологический поперечник мышцы. Для мышц с параллельным ходом волокон физиологический поперечник совпадает с анатомическим.
Отношение МС мышцы к ее физиологическому поперечнику называется абсолютной силой мышцы. Она колеблется в пределах 0,5-1 Н/см2.
Измерение мышечной силы у человека осуществляется при его. произвольном усилии, стремлении максимально сократить необходимые мышцы. Поэтому когда говорят о мышечной силе у человека, речь идет о максимальной произвольной силе (МПС, в спортивной педагогике этому понятию эквивалентно понятие абсолютная сила мышц).
Она зависит от двух групп факторов: мышечных (периферических) и координационных (центрально-нервных).
К мышечным (периферическим) факторам, определяющим МПС, относятся:
1. механические условия действия мышечной тяги - плечо рычага действия мышечной силы и угол приложения этой силы к костным рычагам;
2. длина мышц, так как напряжение мышцы зависит от ее длины;
3. поперечник (толщина) активируемых мышц, так как при прочих равных условиях проявляемая мышечная сила тем больше, чем больше суммарный поперечник произвольно сокращающихся мышц;
4. композиция мышц, т. е. соотношение быстрых и медленных мышечных волокон в. сокращающихся мышцах.
К координационным (центрально-нервным) факторам относится совокупность центрально-нервных координационных механизмов управления мышечным аппаратом - механизмы внутримышечной координации и механизмы межмышечной координации.