
Максимальное потребление кислорода
При плавании с одинаковой скоростью женщины расходуют меньше энергии, чем мужчины, главным образом потому, что у женщин больше плавучесть.
С увеличением скорости плавания потребление О2 возрастает при плавании вольным стилем экспоненциально (примерно пропорционально квадрату скорости), а при плавании брассом и дельфином - линейно, лишь несколько замедляясь при большой скорости (рис. 79). Такой характер зависимости между энергетическими расходами (потреблением О2) и скоростью плавания разными способами объясняется прежде всего особенностями изменения лобового сопротивления и механической эффективности.
Энергетические расходы при плавании брассом и дельфином вдвое больше, чем при плавании вольным стилем.
Наибольшее потребление О2, которое может быть достигнуто при работе только руками или только ногами, составляет соответственно 70-80 и 80-90% от наибольшего его потребления при полноценном плавании. Максимальная скорость плавания при работе руками меньше, чем при работе руками и ногами, что соответственно ведет к более низкому потреблению О2.
Однако при плавании кролем это различие крайне мало, что связано с высокой эффективностью гребков руками.
На дистанции 100 м (50-60 с) примерно 80% энергии обеспечивается анаэробным путем (околомаксимальная анаэробная мощность). С увеличением дистанции возрастает аэробный компонент энергопродукции: на
дистанции 400 м он превышает 50% общей энергопродукции. На дистанциях 800 и 1500 м очень важную роль играют мощность и емкость кислородной системы.
Скорость, начиная с которой содержание молочной кислоты в крови быстро увеличивается (анаэробный лактацидемический порог), соответствует примерно 80% от МПК. Тренированные пловцы способны работать на относительно высоком уровне потребления О2 (60-70% от МПК) без повышения содержания лактата в крови. При максимальной скорости плавания анаэробный гликолиз обеспечивает 50-60% энергии.
Максимальная концентрация лактата в крови у высококвалифицированных спортсменов достигает 18 ммоль/л.
Эффективность плавания крайне низкая.
Даже у высококвалифицированных пловцов она составляет 4-7%.
(Для сравнения: механическая эффективность наземной работы -ходьбы, бега, работы на велоэргометре - 20-30%. Отметим, однако, что при работе на ручном эргометре на суше эффективность также низкая - примерно 10%).
Наибольшая эффективность отмечается при плавании кролем - 6-7% (максимум до 15%), наименьшая - брассом (4-6%).
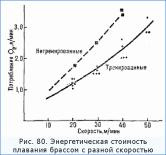
При одинаковой скорости плавания (одним, и тем же способом) тренированный пловец расходует заметно меньше энергии, чем нетренированный (рис. 80).
Эффективность плавания у нетренированного человека может быть в 8 раз меньше, чем у высококвалифицированного пловца. Индивидуальные колебания механической эффективности в плавании значительно больше, чем в таких видах наземной спортивной деятельности, как бег, ходьба, работа на велоэргометре.
Исключительно большие различия в потреблении О2 не только между нетренированными и тренированными людьми, но даже между высокотренированными пловцами указывают прежде всего на сложность плавательной техники. Кроме того, большое значение, как уже отмечалось, имеют размеры и форма тела (определяющие лобовое сопротивление), положение тела в воде, размеры и подвижность весел, создающих движущую силу.
В определенных пределах с увеличением скорости плавания, вплоть до оптимальной, эффективность нарастает. При дальнейшем увеличении скорости она падает.
Оптимальная скорость зависит от способа плавания и техничности пловца. В диапазоне относительно небольших скоростей (0,4- 1,2 м/с) для данного человека энергетическая стоимость проплывания (кролем) 1 км постоянна, т. е. не зависит от скорости плавания.
Пловцы с плохой техникой расходуют больше энергии на единицу дистанции при любой скорости.
Удельный вес девочек и мальчиков вплоть до периода полового созревания заметно не различается. Соответственно и энергетическая стоимость плавания (со скоростью 0,7 м/с) на единицу дистанции с учетом размеров тела у них одинакова. Примерно с 15 лет этот показатель значительно снижается у девушек и повышается у юношей.
На сверхдлинных дистанциях оптимальное соотношение между лобовым сопротивлением и механической эффективностью у женщин более чем компенсирует их сравнительно низкое МПК- Это объясняет определенное преимущество женщин перед мужчинами в плавании на сверхдлинные дистанции. Энергетическая стоимость проплывания 1 км дистанции составляет у нетренированных женщин 250 - 300 ккал, у нетренированных мужчин - 400 - 500 ккал, у спортсменок - 75- 150 ккал, у спортсменов-150 - 200 ккал.
Максимальное потребление кислорода
У нетренированных (в плавании) людей МПК при плавании в среднем на 15-20% ниже, чем в наземных условиях (например, при беге на тредбане). Чем выше тренированность пловца, тем ближе его плавательное МПК (определяемое при плавании) к абсолютному (наземному). У высокотренированных пловцов плавательное МПК в среднем примерно лишь на 6-8% ниже абсолютного, выявленного во время бега в гору на тредбане, и примерно равно МПК при работе на велоэргометре.
У выдающихся пловцов МПК при плавании такое же, как и при беге, или даже немного выше.
Эти данные говорят о высокой специфичности плавательной тренировки, что связано с такими уникальными особенностями плавания, как горизонтальное положение тела в воде (в отличие от обычного вертикального положения при работе в наземных условиях), активация меньшей мышечной массы к преимущественная работа мышц рук и пояса верхних конечностей (в отличие от преобладающей работы мышц ног и туловища при наземных локомоциях).
Следовательно, МПК,. измеряемое в наземных условиях, не может быть полноценно использовано для оценки аэробной работоспособности пловца, а его тренировка, направленная на увеличение максимальной аэробной мощности, должна быть в основном плавательной.
Во время плавания различными способами МПК достигается при неодинаковых скоростях: в брассе - при меньшей скорости, чем в других способах. При одинаковом способе плавания менее тренированные спортсмены достигают своего уровня МПК при более низких скоростях, чем более тренированные пловцы.
Выдающиеся пловцы, особенно стайеры, отличаются высоким МПК - в среднем 5,2 л/мин (4-6 л/мин) при плавании и 5,4 л/мин (4,7-6,4 л/мин) при беге на тредбане, т. е. разница составляет в среднем 5,6%. Соответствующие показатели у женщин - 3,4 л/мин (2,9-3,7 л/мин) и 3,6 л/мин (3,4-4 л/мин).
Относительное беговое МПК (на 1 кг веса тела) у мужчин составляет в среднем 68,6 мл/кг-мин (62,5-76,4), у женщин -55,3 мл/кг-мин (47,8-61,2), что ниже, чем у представителей земных видов спорта, требующих проявления выносливости.
Пловцы обычно весят больше, чем бегуны-стайеры. Поэтому относительное МПК у пловцов меньше, чем у хороших стайеров. Во время плавания вес тела слишком мал и в отличие от наземных локомоций не играет практически никакой роли как фактор нагрузки.
Расход энергии при плавании не пропорционален весу тела, как при беге. Поэтому максимальные аэробные возможности у пловцов лучше оценивать по абсолютному МПК ( л /мин ).
Кислород транспортная система
Большинство физиологических особенностей при плавании обусловлено реакцией организма на пребывание в воде (водную иммерсию), горизонтальное положение тела, давление окружающей среды на тело и преимущественную работу верхними конечностями.

Рис. 81. Общая емкость легких (ОЕЛ), резервный объем вдоха (РОВД), резервный объем выдоха (РОВЫД) и резервный (остаточный) объем (РО) при беге и плавании у 3 испытуемых
л больше остаточного объема.
Для пловцов характерна большая ЖЕЛ: у мужчин-пловцов высокого класса - 5-6,5 л, у женщин - 4-4,5 л, что в среднем на 10-20% больше, чем у людей того же возраста и пола, не занимающихся плаванием. Сила дыхательных мышц и емкость легких у пловцов, определяемые количеством воздуха, выдыхаемого за первую секунду форсированного выдоха, также на 8-15% больше обычных величин.
Давление воды и ее выталкивающая сила обусловливают определенные приспособительные особенности дыхания. Статические размеры легких при погружении тела в воду (без опускания головы) несколько уменьшаются. ЖЕЛ в воде снижается примерно на 8-10%. Частично (около 3%) это связано с увеличением объема крови в грудной клетке (т. е. центрального объема крови) и в некоторой мере (5-7%) с напряжением дыхательной мускулатуры, противодействующей гидростатическому давлению воды.
При плавании ЖЕЛ уменьшается также за счет горизонтального положения тела. Функциональная остаточная емкость становится лишь на 0,5-1,1
В отличие от дыхания в воздушной среде в воде дыхательный объем увеличивается исключительно за счет использования резервного объема вдоха - РОВд (рис. 81). Резервный объем выдоха (РОВыд) уменьшается до 1 л (в условиях воздушной среды до 2,5 л).
Уровень спокойного дыхания смещается в сторону остаточного объема, уменьшая функциональную остаточную емкость. В результате во время дыхания в воде состав альвеолярного воздуха изменяется очень значительно при каждом дыхательном цикле.
Альвеолярная вентиляция при максимальном аэробном плавании (потребление О2 на уровне МПК) выше, чем при максимальной аэробной работе на суше.
Сопротивление току воздуха в дыхательных путях при водной иммерсии в условиях покоя и во время активного плавания возрастает более чем на 50% и требует увеличения активности дыхательных мышц. При плавании кролем дополнительное количество кислорода на каждый литр вдыхаемого воздуха достигает 1,32,8 мл.
Дыхание во время плавания синхронизируется с плавательными (гребковыми) циклами: длительность фазы вдоха уменьшается, а выдох удлиняется и обычно производится под водой (за исключением брасса и плавания на спине), т. е. против большего сопротивления, чем в воздушной среде, - дополнительно примерно на 50-100 мм вод. ст.
Во время плавания с субмаксимальным потреблением О2 легочная вентиляция, дыхательный коэффициент, парциальное напряжение и процент насыщения артериальной крови кислородом связаны с потреблением О2 примерно так же, как и при беге на тредбане или при работе на велоэргометре.
Легочная вентиляция и число гребков в минуту являются линейными функциями скорости плавания, хотя у разных людей имеются довольно значительные вариации в положении и наклоне линий связи между этими переменными. Дыхательный объем в 2-3 л отмечается при частоте гребков 42-73 в минуту. Максимальная легочная вентиляция варьирует от 118 л/мин (ВТР8) у специализирующихся в кроле, брассе и дельфине до 159 л/мин у плавающих на спине.
При плавании на спине частота дыхания доходит до 64 циклов в минуту (примерно два цикла приходится на полный гребковый цикл), а при других способах плавания - до 40.
Вентиляционный эквивалент О2 при максимальном аэробном плавании ниже, чем при аналогичной наземной работе. Причины такой относительной гиповентиляции - особые механические условия: давление воды на грудную клетку, затрудняющее дыхательные экскурсии, зависимость дыхания от ритма плавания (частоты гребковых движений). При одинаковом уровне потребления О2 легочная вентиляция в плавании кролем обычно на 30% меньше, чем в беге или в плавании на спине.
Средние величины легочной вентиляции при максимальном аэробном плавании также ниже, чем при максимальном аэробном беге (на уровне земного МПК). Частота дыхания в плавании ниже, чем в беге.
Несмотря на относительную гиповентиляцию, парциальное напряжение и содержание О2 в артериальной крови при плавании примерно такие же, как и при наземной мышечной деятельности. Хотя альвеолярноартериальный О2-градиент при максимальном аэробном плавании ниже, чем при максимальном аэробном беге, насыщение артериальной крови кислородом составляет около 91 %, т. е. такое же.
Таким образом, легочная вентиляция даже во время максимального аэробного плавания достаточна, чтобы насытить артериальную кровь кислородом до такой же степени, что и во время бега. Следовательно, внешнее дыхание, как и на суше, не ограничивает МПК.
Более низкое МПК при плавании, чем при наземной локомоции, не связано с относительно сниженной вентиляцией.
Сердечно-сосудистая система

Сердечный выброс во время плавания увеличивается почти линейно с ростом потребления О2 (скорости плавания), и при одинаковом субмаксимальном потреблении О2 он примерно такой же, как и при беге или работе на велоэргометре (рис. 82).
Максимальный сердечный выброс у тренированных пловцов во время плавания такой же, как при беге, а у нетренированных пловцов может быть на 25% ниже.
Частота сердечных сокращений во время плавания возрастает линейно с увеличением потребления О2 (скорости плавания); она обычно несколько ниже, чем при беге или работе на велоэргометре с таким же уровнем потребления О2. Это необходимо учитывать, когда ЧСС используется как показатель нагрузки.
Снижение температуры воды уменьшает ЧСС, что компенсируется увеличением систолического объема.
Максимальная ЧСС при плавании также меньше, чем при беге, в среднем на 10-15 уд/мин. У мужчин она составляет в беге около .200 уд/мин, а в плавании - около 185 уд/мин, у женщин соответственно около 200 и 190 уд/мин.
Как и во время работы на суше, во время плавания с одинаковой относительной аэробной нагрузкой (с равным % МПК) ЧСС у тренированных и нетренированных пловцов в среднем одинакова.
Систолический объем растет при переходе от покоя к легкой работе и далее увеличивается с ростом мощности работы (скорости потребления О2). При относительно небольших аэробных нагрузках он достигает определенного уровня, а затем, несмотря на увеличение нагрузки (скорости плавания), вплоть до максимальной, остается неизменным или лишь слегка увеличивается (см. рис.
82).
Горизонтальное положение тела создает благоприятные условия для усиленного венозного возврата и соответственно для большого заполнения сердца во время диастолы. Поэтому при одинаковом субмаксимальном уровне потребления О2 систолический объем во время плавания больше, чем во время работы на велоэргометре, что соответственно ведет к снижению ЧСС во время плавания.
При максимальной аэробной нагрузке в плавании достигается наибольший для данного человека систолический объем. У тренированных пловцов он такой, же, как и при беге, а у нетренированных - ниже,
чем при беге. Как и у представителей других видов спорта, требующих проявления выносливости, систолический объем у пловцов в значительной мере определяется объемом (дилятацией) полостей сердца.
Системная АВР-О2 при субмаксимальном уровне потребления О2 примерно одинакова в плавании и в беге, а при максимальном аэробном плавании несколько меньше по сравнению с максимальным аэробным бегом (соответственно около 15- и 16%).
Содержание О2 в артериальной крови примерно одинаково во время плавания и бега. Максимальная экстракция О2 работающими мышцами из крови также одинакова: минимальное содержание О2 в крови бедренной вены в обоих случаях составляет около 2 об%.
Следовательно, различие в максимальной системной АВР-О2 отражает, по-видимому, особое распределение кровотока при плавании с увеличением кровоснабжения неактивных органов и тканей тела.
Поскольку максимальная АВР-О2 при плавании и беге почти одинакова, сниженное при плавании МПК почти целиком объясняется уменьшенным в воде максимальным сердечным выбросом (из-за снижения максимальной ЧСС). Однако квалифицированные пловцы способны увеличивать свой систолический объем во время плавания, компенсируя сниженную ЧСС и поддерживая. максимальный сердечный выброс.
В значительной мере механизм этого эффекта в усиленном венозном возврате за счет эффективного действия мышечного насоса. У неквалифицированных пловцов этот механизм недостаточно развит.
Среднее артериальное давление при субмаксимальной и максимальной нагрузках в плавании больше, чем в беге, обычно на 10-20%. Это может быть результатом повышенного внешнего (гидростатического.) давления .на тело и увеличения периферического сосудистого сопротивления кровотоку из-за сужения кожных кровеносных сосудов вследствие низкой кожной температуры (26-28). Определенную роль играет также количество участвующей в работе мышечной массы.
Известно, что сокращение небольших групп мышц вызывает более высокий подъем кровяного давления, чем напряжение больших мышечных групп.
При вертикальном положении тела на суше перфузионное давление в сосудах работающих ног повышено за счет гидростатического давления столба крови. Поэтому перфузия крови при беге облегчена по сравнению с горизонтальным положением тела при плавании. Однако повышенное АД во время плавания может вызвать усиление перфузии крови через сосуды работающих мышц, создавая благоприятные условия для снабжения их кислородом .
Локальные (мышечные) факторы
Исключительно важную роль в плавании, как и в других видах спорта, играют функциональные возможности исполнительного мышечного аппарата. Особую роль играют мышцы рук и пояса верхних конечностей, а при брассе - и мышцы ног.
Исследования композиции мышц показали, что у пловцов более высокий процент медленных волокон, чем у неспортсменов (соответственно 74,3 и 46% в дельтовидной мышце и 52,7% и 36,1% в четырехглавой мышце бедра). Аналогичные данные были получены в исследованиях композиции мышц у спортсменок и нетренирован-вых женщин.
У пловцой-спринтеров быстрые волокна составляют 60-65% всех волокон дельтовидной мышцы.
В процессе плавательной тренировки происходит усиление окислительного потенциала быстрых волокон, так что рабочие мышцы почти не имеют быстрых гликолитических волокон (II-В) и содержат практически только быстрые окислительные волокна (II-А) - см. табл. 16.
Наряду с высоким процентом медленных волокон и уровнем их окислительного потенциала преобразование быстрых волокон в быстрые окислительные создает большой аэробный потенциал для рабочих мышц пловца.
К этому следует добавить усиленную капилляризацию рабочих мышц, что наряду с повышением активности ферментов окислительного метаболизма, увеличением количества и размеров митохондрий, содержания миоглобина и другими локальными изменениями ведет к повышению аэробных возможностей этих мышц. Это находит свое отражение в повышении МПК и аэробной работоспособности (выносливости) пловца.
Терморегуляция
Те мпература воды обычно ниже температуры кожи. Вода обладает большой теплоемкостью и теплопроводностью, что в сочетании с конвекцией (движением воды вдоль тела) создает предпосылки для значительных теплопотерь в воде. Если в условиях воздушной среды человек поддерживает тепловой баланс (постоянную температуру тела), несмотря на большие колебания температуры воздуха, то в условиях водной иммерсии для поддержания нормальной температуры тела без его теплоизоляции или усиления теплопродукции необходима температура воды около 33 Самая низкая температура воды, при которой в условиях полного покоя может поддерживаться тепловой баланс (критическая температура воды), варьирует
от 22 (для полных) до 32 (для худых). Быстрая потеря тепла в воде особенно опасна для пловцов-стайеров и ныряльщиков, длительно находящихся в воде.
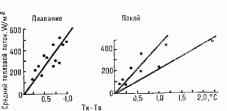
Средний поток тепла от кожи в воду определяется разностью между средней температурой кожи и температурой воды. В покое температура кожи на 1-2 выше температуры воды, а при активном плавании эта разница менее 1 Тем не менее тепло так быстро отводится от поверхности тела в воду, что теплопотери определяются (лимитируются) главным образом тканевой проводимостью, которая, в свою очередь, зависит от разности между температурой ядра и температурой кожи. При этом передача тепла не зависит от скорости плавания (рис.
83).
При температуре воды лишь на 2 ниже нейтральной (33) быстро происходит сужение периферических (кожных и мышечных) сосудов, что увеличивает тканевую изоляцию: уменьшается проведение тепла от ядра тела к коже, т. е. снижаются потери тепла телом. Очень теплая вода обусловливает вазодилятацию и уменьшение тканевой изоляции (увеличение потери тепла). Тканевая изоляция прямо зависит от толщины подкожного жирового слоя. Изменения температуры ядра тела в воде обратно связаны с толщиной подкожного жирового слоя.
Поэтому уменьшение проведения тепла за счет снижения кожного кровотока особенно важно для худых людей. Например, при температуре воды 26 эзофагальная температура у худых пловцов снижается на 0,2, а у полных даже увеличивается на 0,3-0,9.
После погружения в воду с температурой 10 через 10;-20 мин температура тела (измеренная во рту) падает до 32,5, а при зимнем купании- до 30. И здесь большое значение имеют толщина подкожного жирового слоя и степень тренированности к таким условиям.
Так, при температуре воды 16 нетренированный худой мужчина вынужден покинуть ее через 30 мин, когда его ректальная температура снижается до 34,5. Достаточно полный тренированный мужчина в этих же условиях может плавать более 6 ч без изменения ректальной температуры.
Во время плавания около 95% всей энергопродукции превращается в тепло. Как уже отмечалось, плавание увеличивает тканевое проведение тепла, что вызывает его отдачу телом, особенно в прохладной воде. При этом теплоотдача больше, чем теплопродукция.
Соответственно в прохладной воде (ниже' 25) .тело охлаждается более быстро при активном плавании, чем при неподвижном положении.
Реакция кровообращения на движения в воде отражает конфликтные запросы к метаболизму (снабжение рабочих мышц кислородом), с одной стороны, и к нормальной терморегуляции, с другой. Ни одно из этих требований (особенно в холодной воде) полностью не удовлетворяется.
Большие терморегуляторные нагрузки (охлаждение тела) ведут к снижению кровоснабжения мышц из-за усиления кожного кровотока.
Если в результате охлаждения температура ядра тела падает ниже 37, потребление О2 повышается примерно на 0,5 л/мин при любой субмаксимальной скорости плавания. МПК уменьшается на 6-18% по сравнению с МПК при нормальной температуре тела. Повышенное потребление О2 при субмаксимальной скорости плавания и снижение МПК сильно уменьшают работоспособность (выносливость) пловца в условиях пониженной температуры воды (рис.
84).
При интенсивном и непродолжительном плавании в обычных бассейнах с оптимальной температурой воды тепловой баланс организма пловца практически не нарушается. Более того, создаются условия, при которых у пловца относительно меньшая часть сердечного выброса направляется в кожную сеть (как терморегуляторный механизм), чем у бегуна на длинную дистанцию в жарких условиях.
Глава 9. Физиологические особенности спортивной тренировки женщин
Физиологические реакции на физическую нагрузку, а также и механизмы, определяющие функциональные возможности организма и их изменение под влиянием спортивной тренировки, у женщин и мужчин принципиально не различаются Некоторые количественные различия между ними хорошо иллюстрируются соотношением мировых спортивных рекордов. Рекордные результаты у женщин на беговых дистанциях на 813% ниже, чем у мужчин.
В плавании женские рекорды несколько ближе к мужским, чем в беге (разница 610%).
Зависимость функциональных возможностей организма от размеров тела
При сравнении функциональных показателей у женщин и мужчин следует прежде всего учитывать различия в размерах тела. В среднем женщины ниже ростом, чем мужчины.
Даже только из-за этих различий при всех других одинаковых условиях многие функциональные показатели у женщин, в частности их работоспособность, должны отличаться от соответствующих показателей у мужчин.