
ВНУТРЕННЯЯ ОПЕРАЦИОНАЛЬНАЯ АРХИТЕКТОНИКА
Ясно, что на первый план в формировании истинно функциональных систем выступают законы результата и динамической мобилизуемости структур, обеспечивающие быстрое формирование функциональной системы и получение данного результата.
Нам хотелось бы отметить одно важное обстоятельство, которое проходит мимо внимания исследователя. Это свойство внезапной мобилизуемости структурных элементов организма в соответствии с непрерывными функциональными требованиями, которые функция предъявляет к структуре.
Под свойством мобилизуемости мы понимаем возможность моментального построения любых дробных комбинаций, обеспечивающих функциональной системе получение полезного приспособительного результата.
Практически, если бы не было этой потенциальной способности структур к
4. Схема иерархических объединений функциональных систем различной сложности, но построенных на одном пейцмекерном физиологическом образовании, например на возбуждениях пищевого центра. Схема дает возможность сопоставить реальную организацию функциональных систем Z\: 5, : : : б имеющих общий пейцмекер (Рг).
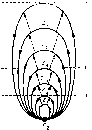
пас-тоящему изолированные функциональные системы организма, можно только с дидактической целью выбрать определенную систему, обеспечивающую какой-то результат па данном уровне иерархии систем (рис. 4).
Так, например, соотношение актина и актомиозина, конечно, составляет по своей операциональной архитектонике вполне очерченную функциональную систему, заканчивающуюся положительным результатом, который можно было бы сформулировать как сокращение мышечной фибриллы. Но такая функциональная система представляется лишь промежуточной между еще более тонкими молекулярными соотношениями протоплазмы мышечной клетки и между движением (например, движением охотника по лесу в поисках дичи), поскольку это движение осуществляется, в конце концов, также с помощью актина и актомиозина.
Но как обширен диапазон, в который включено множество функциональных систем, составляющих эту грандиозную иерархию систем!
Естественно поэтому, что, говоря о составе функциональной системы, мы должны иметь в виду, что каждая функциональная система, взятая для исследования, неизбежно находится где-то между тончайшими молекулярными системами и наиболее высоким уровнем системной организации в виде, например, целого поведенческого акта.
Нетрудно понять, что, раскрыв реальные физиологические механизмы объединения функциональных систем различных уровней, мы приблизились бы к решению проблемы органического объединения анализа и синтеза в самом исследовательском процессе.
В самом деле, сокращение мышечного волокна представилось бы нам в двух аспектах: с одной стороны, это могло бы быть сокращение как процесс вообще сократительных структур, а с другой сокращение, составляющее какую-то очень дробную субсистему, например в спортивном прыжке. Ясно, что мы имеем грандиозное различие в составе этих систем и, может быть, еще более грандиозную задачу определения места компонента в большой системе.
В последние годы вопрос об уровнях организации больших систем особенно подчеркивается рядом советских (Кремянский, Введенов и др.) и зарубежных авторов (Джерард, Браун, Новиков и др.). Наиболее полный обзор этого направления в поисках ннтегративпых закономерностей дан в книге и статьях В. И. Кремянского.
Это направление выступает под различными названиями: интегративные уровни, структурные уровни, иерархия систем и др.
Однако опять-таки в силу того же излишнего теоретизирования ни одни из авторов не дает ничего конструктивного для конкретного исследовательского процесса и не отвечает на основные вопросы иерархии.
В сущности на это указывает и употребление термина уровни, который находится в абсолютном противоречии с понятием система. Главное же то, что пи в одной концепции уровни не обладают какой-либо функциональной архитектоникой и, следовательно, как способ соединения уровней, так и механизмы, удерживающиее единство всей архитектуры целого, естественно, не могут быть найдены.
Все сказанное выше с совершенной очевидностью убеждает нас в том, что перед исследователем стоят по крайней мере два кардинальных вопроса, не решив которые он не может надеяться на понимание тонких механизмов сложных функциональных систем организма.
Несколько лет назад эти вопросы были сформулированы нами в следующем виде.
1. Различается ли чем-либо принципиально архитектура функционирования как у весьма элементарных, так и у сложных субсистем? Иначе говоря, функционируют ли системы всех уровней по одной и той же архитектуре, которая характерна для функциональной системы вообще, или эти архитектуры чем-то отличаются друг от друга?
2. Какими конкретными механизмами соединяются между собой субсистемы при образовании суперсистемы? Учитывая наличие в функциональной системе определенного количества специфических для нее узловых механизмов, вопрос можно поставить более конкретно: какими именно узловыми механизмами своей архитектуры соединяются субсистемы, чтобы образовать суперсистему?
Не объединяются ли но этому же принципу и большие системы промышленных предприятий? Мы предоставляем право судить об этом соответствующим специалистам. Но если бы это было так, перед нами встал бы вопрос о замечательной
гармонии в организации всех тех систем, где результат является решающим фактором системообразования.
Мы допускаем, что при анализе такого ответственного вопроса, как объединение субсистем в суперсистему, могут возникнуть другие вопросы, однако несомненно одно, что без решения двух поставленных выше вопросов нельзя надеяться на решение всей проблемы иерархии систем в целом. Они принципиальны по своей сути.
Отвечая на первый вопрос, нужно исходить из того вывода, к которому мы пришли при формулировке самого понятия система. Центральным моментом для системы является результат, так как любой комплекс и любое множество становятся системой только благодаря результату.
Вместе с тем система не может быть стабильной, если сам результат своими существенными параметрами не влияет на систему обратной афферентацией. А если это так, то любая система, какой бы значительной она ни была в иерархическом ряду, должна подчиняться этим правилам.
Все эти соображения приводят нас к окончательному и фундаментальному выводу о составе иерархии: все функциональные системы независимо от уровня своей организации и от количества составляющих их компонентов имеют принципиально одну и ту же функциональную архитектуру, в которой результат является доминирующим фактором, стабилизирующим организацию систем.
Решение второго из поставленных выше вопросов облегчается принятием положения, что архитектуры систем принципиально тождественны.
Если допустить, что какие-то субсистемы соединяются между собой, вступают в контакт с помощью каких-то промежуточных механизмов, ведущих к получению полезного результата, то сразу же будет видно, что такое допущение не может быть сделано. Тогда какие-то субсистемы не смогут развить своего основного функционального смысла, т. е. получения результата, и, таким образом, сама система не может быть названа системой.
Поэтому наиболее вероятно, что именно полезный результат системы, какой бы малой она ни была, представляет тот реальный вклад, который она может сделать при образовании суперсистемы, или большой системы.
Отсюда следует, что при образовании иерархии систем всякий более низкий уровень систем должен как-то организовать контакт результатов, что и может составить следующий, более высокий уровень систем и т. д. Очевидно, организм формирует свои системы именно таким образом, и только при этом возможно организовать системы с обширным количеством компонентов. Естественно, что в этом случае иерархия систем превращается в иерархию результатов каждой из субсистем предыдущего уровня.
Другой важный вопрос, возникающий при образовании иерархии систем, состоит в следующем: как действует эта субординированная иерархия, когда ей надо выступать как целое?
Хороший пример представляет соотношение уровня кровяного давления и какого-либо эмоционального состояния, возникшего под влиянием внешних воздействий. Мы знаем, что при возникновении эмоции, например страха, происходит быстрый подъем кровяного давления, что имеет несомненное приспособительное значение.
Но в то же время мы знаем, что постоянный уровень кровяного давления представляет собой результат самостоятельной разветвленной функциональной системы, независимой от эмоционального разряда (П. К. Анохин, 1960). Как эмоциональный разряд, возникший по внешнему поводу, находит
доступ к функциональной системе кровообращения? На какие компоненты этой более низко организованной системы действует эмоциональный разряд?
Кора головного мозга и вызванная ею эмоция заинтересованы в поддержании высокого уровня кровоснабжения и метаболических процессов в условиях стрессового состояния целого организма. Поэтому ясно, что эмоциональный разряд должен оказать свое действие на результат функциональной системы на уровень кровяного давления.
Но этот уровень физическая величина.
Таким образом, эмоциональный разряд должен действовать прежде всего на эфферентные механизмы, определяющие уровень кровяного давления. Следовательно, в нисходящем направлении эмоциональный разряд должен подействовать на эфферентное звено системы, определяющее уровень давления, т. е. на сосудосуживающий центр.
Обращает на себя внимание один весьма интересный факт: в этом случае уровень кровяного давления не зависит от того афферентного синтеза, который производится ежесекундно сосудосуживающим центром на основе барорецепторной афферентной сигнализации. В случае эмоционального разряда возбуждение суперсистемы прямо занимает эфферентные пути субсистемы и устанавливает нужный уровень кровяного давления, минуя афферентный синтез хозяина субсистемы.
На долю же этой последней остается лишь сопротивляться при помощи барорецепторов чрезмерному давлению, оказываемому на ее сосуды...
Подводя итог сказанному, следует заметить, что главной чертой каждой функциональной системы является ее динамичность. Структурные образования, составляющие функциональные системы, обладают исключительной
мобилизуемостью. Именно это свойство систем и дает им возможность быть пластичными, внезапно менять свою архитектуру в поисках запрограммированного полезного результата.
ВНУТРЕННЯЯ ОПЕРАЦИОНАЛЬНАЯ АРХИТЕКТОНИКА ФУНКЦИОНАЛЬНОЙ СИСТЕМЫ
Не будет преувеличением сказать, что трудность развития системного подхода вообще и общей теории систем Берталанфи в частности состоит именно в том, что обсуждение ведется на уровне глобальных свойств системы, так сказать обсуждение системы черного ящика. Подавляющее большинство исследователей не делают попытки проникнуть во внутреннюю архитектонику системы и дать сравнительную оценку специфических свойств ее внутренних механизмов.
При таком подходе обсуждаемая система всегда выглядит как нечто гомогенное, в котором клетки одинаковы, все компоненты равноценны и все механизмы разнозначны.
В действительности же дело обстоит совсем наоборот. Функциональная система всегда гетерогенна. Она всегда состоит из определенного количества узловых механизмов, каждый из которых занимает свое собственное место и является специфическим для всего процесса формирования функциональной системы. Становится очевидным, что, не вскрыв этих своеобразных механизмов, составляющих внутреннюю операциональную архитектонику системы, мы не приблизимся к самой решающей цели системного подхода вообще обеспечению органического единства в исследовательском процессе системного уровня функционирования с индивидуальной характеристикой каждого дробного элемента или механизма, принимающего участие в этом функционировании (рис.
5).
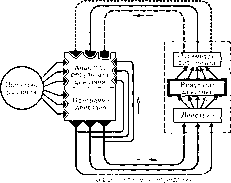
таки он не указывает самого важного: на основе какого же критерия должна быть построена логика механизма и должно быть произведено упрощение множества. Теория функциональной системы решает этот вопрос четко и обоснованно.
Центральным критерием упрощения множества является результат системы, который, как мы видели, предъявляет решающие требования к определенным степеням свободы компонентов системы. С точки зрения теории функциональной системы, улучшенная логика механизма есть не что иное, как внутренняя операциональная архитектоника системы, отвечающая па все требования тончайшего физиологического анализа механизмов системы до молекулярного уровня включительно.
Ниже мы приводим узловые специфические механизмы, представляющие собой внутреннюю архитектонику системы и вместе с тем обеспечивающие объединение в одной исходной концепции и высшего синтеза и тончайшего анализа.
АФФЕРЕНТНЫЙ синтез
Как мы уже установили в начале раздела, для функциональной системы характерно то, что вопрос, какой результат должен быть получен, решается внутри системы и па основе ее закономерных механизмов. Это обстоятельство радикально отличает биосистему от самых сложных машинных устройств автоматической регуляции. Практически для всех машин цель поставлена за пределами машины и для нее допускается лишь некоторая способность самоорганизации в процессе получения запрограммированного не ею результата. Биосистема даже очень простой иерархии сама, на основе своих внутренних процессов, принимает решение о том, какой результат нужен в данный момент ее приспособительной деятельности.
Вопрос этот решается именно в стадии афферентного синтеза.
Поскольку этот процесс много раз описывался нами в различных публикациях, мы дадим здесь лишь краткую формулировку его особенностей. Прежде всего вопрос о его составе.
Какие именно афференты синтезируются в этой стадии?
Мы предложили четыре решающих компонента афферентного синтеза, которые должны быть подвергнуты одновременной обработке с одновременным взаимодействием па уровне отдельных нейронов: доминирующая на данный момент мотивация, обстановочная афферентация, также соответствующая данному моменту, пусковая афферентация и, наконец, память (рис. 6).
Основным условием афферентного синтеза является одновременная встреча всех четырех участников этой стадии функциональной системы. Микроэлектродпын и микрохимический анализы и другие формы аналитического исследования нейрона в момент встречи на нем всех упомянутых выше типов возбуждений показали, что этот процесс поддерживается и облегчается рядом динамических процессов нервной
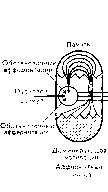
исследовательской реакции; III сенсорно-биологическая конвергенция, благодаря которой различные сенсорные возбуждения имеют возможность вступать в контакт с биологическими состояниями организма, широко генерализованными по огромному количеству корковых и подкорковых нейронов; IV аксонально-сенсорно-биологическая конвергенция, благодаря которой на одном и том же нейроне или на комплексе нейронов создается возможность встречи не только сенсорных, биологических, интрацентральных возбуждений, но и тех возбуждений, которые выходят на конечный нейрон в виде эфферентных. Эти возбуждения отходят по коллатеральным ответвлениям от основного аксона. Такая конвергенция возбуждений с полифункциональным содержанием представляет собой подлинный интегративный блок. системы.
К ним относятся прежде всего восходящая активация, вызванная ориентировочно-исследовательской реакцией, как правило, сопутствующая афферентному синтезу и предшествующая принятию решения, процесс корково-подкорковой реверберации и процесс центробежного повышения возбудимости вовлеченных в афферентный синтез рецепторов.
Своеобразие состоит в том, что этот синтетический процесс, если его отнести к масштабам нейрона, совершается на основе центральной закономерности пнтегратнвяой деятельности мозга, именно на основе конвергенции возбуждений на одном и том же нейроне.
Нам хотелось бы обратить внимание на следующее: одновременность обработки всех четырех типов возбуждений в стадии афферентного синтеза основана на том, что каждый из них приобретает свои особенные физиологические свойства именно в процессе симультапной обработки. Именно здесь происходит освобождение нейрона от избыточных степеней свободы благодаря приходу к нему именно тех, а не других возбуждений.
В связи с этим уместно отметить, что существующая в нейрофизиологии тенденция изучать и рассматривать такие компоненты афферентного
Синтеза, как мотивация, память, стимул и т. п., в качестве отдельных, самодовлеющих проблем неверна и малоэффективна.
Возьмем, например, проблему памяти. Взятая в отдельности, сама по себе, как самодовлеющая проблема, она уже показала, что может повести мысль исследователя совершенно в другую сторону. Действительно, в каком аспекте сейчас развивается проблема памяти?
Подавляющее большинство исследователей проблемы памяти все внимание сосредоточивают на моменте фиксации пережитого опыта живой системы. Это, несомненно, важная сторона памяти, но совсем по-другому выглядит весь вопрос о ней, как только мы будем ее рассматривать как один из компонентов, органически включенных в проблему принятия решения.
Здесь сразу же центр событий перемещается с фиксации опыта на динамическое извлечение этого опыта из молекулярных агрегатов большой системы.
Этот процесс извлечения из памяти является еще более поразительным, если вспомнить, с какой легкостью он ежесекундно и безошибочно помогает принять нужное решение в конце афферентного синтеза. Обратим внимание, с какой легкостью мы извлекаем из памяти самые тончайшие нюансы нашей мысли, разговора и всего того, что было накоплено за всю нашу жизнь.
Совершенно очевидно, что эта чудесная способность памяти быть готовой ежесекундно отдать то, что было накоплено за много лет и что требуется в данной стадии афферентного синтеза, не может быть изучена иначе, как на основе полного контакта и взаимодействия всех четырех компонентов афферентного синтеза.
Значительное облегчение при решении проблемы афферешного синтеза нам принесла разработка концепции об интегративной деятельности нейрона. Она дала возможность сформулировать положение, на основе которого строится динамическое участие памяти в афферентном синтезе: извлечение прошлого опыта из памяти происходит по той же нейрохимической трассе, по которой он был зафиксирован в момент приобретения опыта (П.
К. Анохин, 1974).
Таким образом, афферентный синтез, приводящий организм к решению вопроса, какой именно результат должен быть получен в данный момент, обеспечивает постановку цели, достижению которой и будет посвящена вся дальнейшая логика системы.
Нетрудно видеть, что афферентный синтез, являющийся абсолютно необходимым этапом формирования функциональной системы, содержит все необходимое для постановки цели, которая так долго пугала исследователя-материалиста и так долго находилась в безраздельном владении идеализма.
Все детали афферентного синтеза в пашей лаборатории изучаются на протяжении многих лет с помощью экспериментально-физиологических методик. Так, например, неразделимое единство пусковой и обстановочной афферентации было показано на основе опытов с .жстирпацией лобных отделов коры собаки и последующим изучением реакции выбора на различные условные раздражители (А.
И. Шумилина).
В последнее время были подробно изучены различные виды восходя тих активирующих влияний, которые своеобразно помогают афферентно му синтезу. Здесь прежде всего надо указать на чрезвычайное увеличение дискриминационной способности коркового нейрона к частоте импуль-саций под влиянием мотивационных раздражений гипоталамуса (С.
Н. Хаютин, 1971). Другой важный механизм, выявляющийся при раздражении лимбической системы, это перевод нейронов из категории мономодальных в мультиконвергентную категорию
(В. А. Макаров, 1970).
Легко видеть, что уже только эти два механизма могут значительно повысить информационную емкость нейрона и, следовательно, вместе с другими описанными выше механизмами сделать особенно эффективной стадию афферентного синтеза. К разряду механизмов, облегчающих афферентный синтез, надо отнести также детекторные свойства периферических рецепторов и побочное вытормаживание шумовых процессов при прохождении афферентной информации по центральной нервной системе.
Особенно подробно (до микроэлектродной и до микрохимической методик включительно) была изучена роль доминирующей мотивации в установлении синтетических взаимодействий в этой стадии на поверхности одиночного нейрона. Само собой разумеется, что возможность такого тонкого анализа отдельных процессов, составляющих систему, появилась только тогда, когда была выработана улучшенная логика механизма, т. е. внутренняя операциональная архитектоника системы.
В управленческом деле, как и в биологических системах, также следует поставить вопрос, какой результат должен быть получен системой. Иначе говоря, совокупность всех предварительных расчетов и соображений должна составить (например, в торговом деле) основу афферентного синтеза, приводящего к решению и выбору действия с наилучшим результатом.
Так, например, Ханике (1969), касаясь проблемы управления, пишет: Цель заключается в том, чтобы обеспечить принятие решений на основе систематического анализа.
Насколько отчетливо именно в проблеме управления предприятиями встают вопросы о необходимости афферентного синтеза, совершенно схожие с биологическими проблемами, видно из того удивительного совпадения, которое получилось в аргументации Ханике и нашей собственной по поводу роли афферентного синтеза.
Для выявления наличия широкого афферентного синтеза у человека перед принятием решения рассмотрим пример, когда пешеход переходит улицу. Перед тем как принять решение о переходе, человек должен тщательно оценить довольно большое количество компонентов этого афферентного синтеза (число машин, скорость движения, ширину улицы, свои силы и др.).
Аргументируя наличие этой стадии в системной деятельности человека, Ханике также приводит именно этот пример, перечисляя все параметры, которые должны быть учтены для принятия решения о переходе.
ПРИНЯТИЕ РЕШЕНИЯ
Принятие решения один из самых интересных моментов в развертывании системных процессов. Тот факт, что само понятие принятия решения долгое время изгонялось так называемой чистой наукой из области научных исследований, лучшее доказательство того, что оно происходит на основе весьма комплексных процессов мозга.
В настоящее время, как видно из предыдущего изложения, теория функциональной системы сделала принятие решения полноценным участником объективного процесса формирования системы, критического процесса, благодаря которому афферентная оценка всех условий завершается доступным исследованию механизмом. Афферентный синтез, подчиняясь доминирующей в данный момент мотивации и под коррекцией памяти, ведет такой подбор возможных степеней свободы, при котором возбуждения избирательно направляются к мышцам, совершающим нужное действие.
Возникает весьма существенный для всей нашей концепции вопрос, когда и как происходит принятие решения о получении именно того, а не другого результата.
Последние данные сотрудников нашей лаборатории (А. И. Шумилина, В. Б. Швырков) заставляют думать, что оценка возможных результатов при данной доминирующей мотивации происходит уже в стадии афферентного синтеза.
Однако эти результаты не получаются реально, а следовательно, их оценка происходит при помощи какого-то пока не изученного нами механизма. То же, что происходит в принятии решения, является уже результатом выбора на основе длительной оценки различных, внутренне (!) формирующихся результатов.
Иначе говоря, любое принятие решения, после того как закончится афферентный синтез, является выбором наиболее подходящих степеней свободы в тех компонентах, которые должны составить рабочую часть системы. В свою очередь эти оставшиеся степени свободы дают возможность экономно осуществить именно то действие, которое должно привести к запрограммированному результату.
Как происходит это освобождение от избыточных степеней свободы? Почему момент принятия решения часто имеет характер внезапной интуиции?
В настоящее время мы еще не можем ответить на эти вопросы, но выработанный подход к ним дает полную гарантию того, что принятие решения в биологических системах с большой и малой иерархией является вполне анализируемым и доступным для объективной науки феноменом. Прежние опыты А. И. Шумилиной, которая производила удаление лобных отделов у собаки, находящейся в условиях активного выбора одной из сторон станка, убедительно показали, что в этот момент, т. е. в момент принятия решения, вся обрабатываемая информация интегрируется именно в области лобных отделов, откуда и идет команда к подбору наиболее оптимальных аппаратов поведения.
Интересно отметить, что, являясь наиболее требовательным к объему афферентной информации, процесс принятия решения страдает в первую очередь после различных вмешательств в центральную нервную систему. Весьма доказательной иллюстрацией являются результаты опытов Gambaryan (1971). Разрушая некоторые подкорковые аппараты (бледное тело и Др.), он нашел, что прежде всего длительно страдает активный выбор стороны станка, на которой должно быть сделано пищевое подкрепление.
Наоборот, казалось бы, более сложный произвольный акт нажатие педали для подачи корма остается неразрушенным или мало пострадавшим.
Попытаемся понять сам момент принятия вполне определенного решения с точки зрения общефизиологических закономерностей. Его механизмы станут ясными, если мы представим себе на минуту, какое количество возможных действий может сделать человек, употребляя хотя бы только один мышечный аппарат. Мы уже знаем, что вся эфферентная часть организма должна обладать свойством экстренной мобилизуемости.
Малейшего изменения в распределении мышечных усилий достаточно для того, чтобы было произведено вполне целенаправленное движение конечности или целого организма.
Принципиально таких возможностей движения у организма существует необозримое множество вероятно, не меньше, чем количества комбинаций взаимодействия па площадке с 400 лампочками. Вместе с тем мы знаем, что в каждом
отдельном случае, в соответствии с принятым решением, возбуждения выходят на совершенно определенные мышечные группы. Иначе говоря, происходит очень быстрое отбрасывание всех тех степеней свободы в наших движениях, которые не соответствовали бы принятому решению. Таким образом, сама проблема реализации принятого решения в физиологическом плане должна объяснить два важных вопроса:
1. 'Какие механизмы производят отбор нужных в данный момент степеней свободы и вытормаживают все те степени свободы на моторных нейронах и мышечных аппаратах, которые не имеют отношения к получению данного результата.
2. Где может быть преимущественная локализация этого в высшей степени конденсированного процесса, который одновременно обрабатывает на основе доминирующей мотивации всю прошедшую в мозг аффеферентную информацию, производит непрерывное сопоставление этих результатов с прошлым опытом и, наконец, переводит результаты этой обработки на эфферентные пути, точно соответствующие распределению возбуждений для совершения нужного акта, обеспечивающего получение нужных результатов?
С постановкой этих двух вопросов мы переходим в область, абсолютно новую для нейрофизиологии и потому, естественно, совсем не разработанную. Насколько интересны те новые вопросы исследования, которые при этом возникают, показывает, например, роль доминирующей мотивации в отборе необходимой в данный момент афферентной информации на уровне коры, на ее нейронах.
Микрофизиологическое исследование нейронов вентро-медиального гипоталамуса показало, что уровень их возбудимости зависит от начального возбуждения латерального ядра гипоталамуса голодной кровью.
Дозируя это возбуждение в натуральных условиях сроками голодания (14 дня), мы показали, что количество доминирующего возбуждения меняется на соответствующих синапсах корковых нейронов и в связи с этим значительно меняется интегрирующее влияние доминирующей мотивации на соотношение других синаптических возбуждений данных кортикальных нейронов. Эти исследования дают нам лишнее доказательство того, что системный подход радикально влияет на постановку вопроса каждого конкретного исследования.
Есть основание предполагать, что этот механизм сосредоточен в наиболее интегрирующих и компактных структурах головного мозга. Однако вопрос физиологической интерпретации принятия решения, возникший благодаря системному подходу к предмету, должен быть фактически разработан заново.
ФОРМИРОВАНИЕ АКЦЕПТОРА РЕЗУЛЬТАТОВ ДЕЙСТВИЯ
Аппарат акцептора результатов действия один из самых интересных в деятельности мозга и практически так же универсален и вездесущ, как обратная афферентация и афферентный синтез. Формирование этого механизма нарушает устоявшееся представление о поступательном ходе возбуждений по центральной нервной системе согласно рефлекторному процессу.
Акцептор результатов действия на основе многостороннего механизма афферентного синтеза не является выражением последовательного развития всей цепи явлений поведенческого акта. Он предвосхищает афферентные свойства того результата, который должен быть получен в
соответствии с принятым решением, и, следовательно, опережает ход событий в отношениях между организмом и внешним миром.
Акцептор результатов действия является весьма сложным аппаратом. По сути дела он должен сформировать какие-то тонкие нервные механизмы, которые позволяют не только прогнозировать признаки необходимого в данный момент результата, но и сличать их с параметрами реального результата, информация о которых приходит к акцептору результатов действия благодаря обратной афферентации.
Именно этот аппарат дает единственную возможность организму исправить ошибку поведения или довести несовершенные поведенческие акты до совершенных. Здесь следует также подчеркнуть, что различного рода поиски и компенсации также могут повести к полезному результату через такого рода оценку обратной афферентации. Циркуляторное развитие этих возбуждений при узнавании и поиске может быть столь быстрым, что каждый блок этой функции, состоящей из компонентов: результат обратная афферентация сличение и оценка реальных результатов в акцепторе результатов действия коррекция новый результат и т. д., может развиться буквально в доли секунды.
Особенно быстро этот процесс протекает в условиях сканирования и слежения.
Факт организации этого аппарата непосредственно после принятия решения был доказан в нашей и других лабораториях как в обычном эксперименте с условными рефлексами, так и при помощи тонких электрофизиологических приемов.
Возможно, сам момент встречи опережающего комплекса возбуждений с информацией о реально полученных результатах происходит на основе каких-то весьма тонких признаков возбуждения вообще, допускающих количественное и композиционное сличения.
Одна из интересных форм сличения в акцепторе результатов действия, хотя ее автор и не был знаком с общей архитектурой функциональной системы, была недавно представлена японским ученым Suga (1964). Он изучал получение обратного, отраженного от объекта сигнала при ультразвуковой локации у летучих мышей. Оказалось, что при посылке ультразвукового поискового сигнала в пространство в мозге летучей мыши формируется комплекс из возбужденных клеток с различными латентными периодами, периодом торможения и частотной конфигурацией.
Именно эти комплексы в зависимости от фазы, в которой происходит встреча их с отраженным от объекта ультразвуковым сигналом, решают вопрос о расстоянии, на котором находится преследуемый объект.
Конечно, этот аппарат оценки результата действия сравнительно простой, но он подсказывает нам, какими возможностями и параметрами нервное возбуждение решает проблему сличения. В более сложных формах результата, например при очинке карандаша, этот аппарат, включающий несколько сенсорных компонентов, является, несомненно, более сложным.
Наши последние исследования по составу акцептора результатов действия как комплекса разнородных возбуждений показали, что эти средства ' сличения могут быть очень различны.
Что может входить в состав этого аппарата? Совершенно очевидно, что существенные признаки будущего результата динамически формируются благодаря многосторонним процессам афферентного синтеза с извлечением из памяти прошлого жизненного опыта и его результата.
Некоторые последние данные заставляют, однако, думать, что в стадии
афферентного синтеза складывается несколько возможных результатов, по они не выходят на эфферентные пути и поэтому не реализуются, Решение же совершается после того, как произведен выбор наиболее адекватного результата по отношению к данной доминирующей мотивации.
Этот комплекс возбуждений в подлинном смысле слова афферентная модель будущего результата, и именно эта модель, являясь эталоном оценки обратных афферентаций, должна направлять активность человека и животных вплоть до получения запрограммированного результата.
В последнее время были получены дополнительные сведения о том, что в этот нервный комплекс, обладающий высокой степенью мультикон-вергентного взаимодействия, приходит еще одно возбуждение, совершенно иной, не афферентной, а уже эфферентной природы. Речь идет о коллатеральных ответвлениях пирамидного тракта, отводящих ко многим межуточным нейронам копии тех эфферентных посылок, которые выходят на пирамидный тракт.
Интересно, что эти эфферентные возбуждения конвергируют на те же межуточные нейроны сенсомоторной области, куда поступают и все те афферентные возбуждения, которые могут составить параметры реального результата.
Таким образом, момент принятия решения и начала выхода рабочих эфферентных возбуждений из мозга сопровождается формированием обширного комплекса возбуждений, состоящего из афферентных признаков будущего результата и из коллатеральной копии эфферентных возбуждений, вышедших на периферию по пирамидному тракту к рабочим аппаратам. В зависимости от интервала между постановкой цели и ее реализацией к этому же комплексу возбуждений через определенное время приходят возбуждения и от реальных параметров полученного результата. Самый процесс оценки полученного реального результата осуществляется из сличения прогнозированных параметров и параметров реально полученного результата. Именно здесь, в этом пункте, осуществляется таинство оценки полученного результата, которое, как мы видели на примере с летучей мышью, производится специальным динамически формирующимся аппаратом.
Оценка же и ее результат определяют дальнейшее поведение организма. Если результат соответствует прогнозированному, то организм переходит к следующему этапу поведенческого континуума.
Если же результат не соответствует прогнозу, то в аппарате сличения возникает рассогласование, активирующее ориентировочно-исследовательскую реакцию, которая, поднимая ассоциативные возможности мозга на высокий уровень, тем самым помогает активному подбору дополнительной информации. Все эти процессы поведения акцептора действия достаточно хорошо изучены в нашей лаборатории.
Так, например, было показано, что в момент выхода из коры головного мозга пирамидное возбуждение отдает копию эфферентного возбуждения не только в комплексы оценки результата, т. е. в кору мозга, но и в ретикулярную формацию. Ретикулярная формация в обратном кортикоцетальном направлении имеет возможность оказать дополнительное энергетическое воздействие на те циркулярные возбуждения (круги ожидания), которые должны удержать свою активность до момента прихода информации о получении полезного результата.
Как мы знаем, в некоторых случаях комплекс акцептора результатов действия должен очень долго быть в напряженном состоянии, прежде чем будет получен реальный запрограммированный результат.
ЗАКЛЮЧЕНИЕ
Большая часть раздела посвящена тем разграничительным линиям, которые отделяют общее движение за системный подход от разработанной в нашей лаборатории теории функциональной системы, имеющей и иное начало, и иной путь отработки
принципиальных позиций и понятий.
В одной из своих работ Берталанфи пишет, что идея организменного, или целостного, подхода родилась у пего очень рано, кажется, в 1937 г., однако
интеллектуальный климат того времени не был подходящим для восприятия
организменной идеи, которую он предложил. Действительно, интеллектуальный климат того времени был воспитан па подражании законам физики и механики, и потому считалось научным только то, что могло быть переведено на принципы и понятия этих наук.
И только в 1950 г. Берталанфи решился сделать официальные шаги в защиту своей идеи, ибо в это время, т. е. в послевоенные годы, резко изменился сам
интеллектуальный климат. Весьма быстро и настоятельно проявился интерес к абстрактным формулам, моделям, синтетическим понятиям и вообще к интердисциплипарным взаимоотношениям между учеными, и только в этот период стало возможным говорить об общей теории систем.
Теория функциональной системы претерпела также немало изменений с момента ее первой формулировки, однако условия ее возникновения и самые первые шаги в ее развитии принципиально отличались от путей развития общей теории систем. Как концепция она зародилась примерно в 19321933 гг. и была сформулирована в достаточно развитой форме уже в 1935 г. в сборнике Проблема центра и периферии в физиологии нервной деятельности.
Однако в первых же формулировках функциональная система как интегративное образование организма включала все те компоненты, которые характерны для нее и сейчас, в том числе и обратную, или санкционирующую, афферентацию. В отличие от общей теории систем, которая до сих пор пребывает в стадии исканий конкретных и полезных путей, теория функциональной системы в первые же годы после ее формулировки явилась для нашей лаборатории толчком к бурному развитию именно конкретной исследовательской работы, которая нашла в функциональной системе совершенно очевидный конструктивный стимул к формулировке новых задач исследования.
Эта особенность теории функциональной системы как конкретной конструктивной теории резко отличает ее от судьбы общей теории систем, которая даже к настоящему времени практически еще не имеет связи с конкретной исследовательской работой.
Достаточно указать, что уже в 1937 г. была опубликована кардинальная для наших исследований работа Функциональная система как основа интеграции нервных процессов в эмбриогенезе. Фактически это было зарождением той эволюционной концепции, которая в 1945 г. была сформулирована как теория системогенеза.
Очень часто задают вопрос, к каким принципиальным обобщениям привели исследования, проведенные на основе теории функциональной системы. Системогенез может служить самым демонстративным примером конструктивной продуктивности теории функциональной системы как методологического инструмента, помогающего поставить новые вопросы исследования.
Системогенез приводит к существенным изменениям имевшихся представлений о путях эмбрионального развития. Это стало возможным только потому, что теория функциональной системы оказалась конструктивным принципом, подсказывающим положительно в каждой области новый подход, новые трактовки и новые формы экспериментирования.
Мы не можем здесь перечислять все те направления исследований в различных областях биологии и физиологии, которые получили совершенно новое освещение при использовании теории функциональной системы как рабочего принципа в повседневном исследовании.
[Казалось бы, общая теория систем в соответствии с ее весьма радикальными и даже эволюционными стремлениями в борьбе с механистическим аналитизмом должна была бы вызвать немедленное преобразование самой логики научного исследования и, несомненно, приобрести всеобщую симпатию реально мыслящих ученых. Однако этого не произошло.
Почему же этого не случалось?
В этом разделе мы пытались выявить причины этого парадоксального явления. Нам кажется, ясным, что имеются две причины того ничтожного результата, который был получен при обсуждении общей теории систем.
Первая состоит в том, что авторы ее пошли по неправильному пути как в поисках понятия системы, так и в общей тактике ее разработки.
Как можно было видеть, при всякой попытке сформулировать само понятие системы, т. е. дать принципиальное кредо этому новому интересному направлению, положительно все исследователи попадают в заколдованный круг традиционных понятий. Здесь непрерывно цитируются целостность, организменность,
взаимодействия, организованная сложность, упорядоченное множество и другие подобные термины, которые становятся даже центральными критериями понятия системы. Ясно, что все эти термины по самой своей сути являются лишь вариациями понятия целостности.
Именно поэтому, не выходя за пределы понятийного поля целостности, все эти определения не дают какого-либо существенного скачка как в понимании системы, так и в конкретной разработке ее.
Между тем главный смысл системного подхода состоит именно в том, что любая деталь наблюдения или экспериментирования должна быть неизбежно вписана в какой-то из узловых механизмов внутренней архитектоники системы. Практически никакая новая научная тема не могла бы быть сформулирована вне конкретной внутренней архитектоники системы, где эта тема только и может получить широкий познавательный смысл.
Точно так же и трактовка полученных результатов даст наибольший эффект, если она будет построена па основе системных механизмов.
Вторая причина состоит в том, что интеллектуальный климат для принятия системного воззрения действительно изменился. Однако подавляющее большинство ученых с большим трудом отказываются от устоявшихся традиций рассматривать все научное накопление в аналитическом аспекте.
Такой переход не простая перемена названий и выражений, как поначалу думали некоторые исследователи; он требует радикального изменения caMpix принципов подхода к элементарным процессам и общей тактике исследования.
Действительно, для исследователя, имеющего в руках теорию функциональной системы как методологический инструмент экспериментирования, не может быть возбуждения вообще, афферентации вообще, мотивации вообще и даже памяти
вообще. Конструктивная роль этих привычных понятий выявляется благодаря их положению в том или ином качественно своеобразном механизме внутренней архитектоники функциональной системы.
Возьмем для примера зрительную афферентацию. Она обычно определяется как сенсорная модальность, и с точки зрения этой аналитической характеристики специфической чертой этой афферентации будет именно ее оптический характер.
Но так ли обстоит дело, если оценивать зрительную афферентацию по ее месту в архитектонике функциональной системы?
Зрительная афферентацня может быть пусковой афферентацией, как это, например, наблюдается в случае условного зрительного стимула. Но та же зрительная афферентация в других условиях может составлять и обстановочную афферентацию, определяющую совсем другой механизм системы предпусковую интеграцию нервных процессов.
И даже больше того, зрительная афферентация может иметь еще третью и совершенно иную функциональную значимость: она может участвовать в оценке полученного системой результата. Таким образом, сформулировав внутреннюю операциональную архитектонику системы, мы тем самым изменили подход к привычным для нас физиологическим понятиям и процессам.
Еще более заметной становится роль функциональной системы, когда надо, так сказать, анатомировать какое-то сложное явление в работе мозга животных или человека. Так, например, оценка изменения психической деятельности человека после операции на мозге производится обычно при помощи стандартных тестов, а в последние годы при помощи оценки различных электрофизиологических показателей в работе мозга (ЭЭГ, вызванного потенциала, активности нейронов).
Функциональная система вносит определенный порядок и логическую последовательность в эту оценку. Можно задать вопрос, изменился ли афферентный синтез у данного больного при выработке определенного решения. Если он изменился, то больной не может принять адекватного решения. Тогда сейчас же возникает следующий вопрос: какой из компонентов и какой из нейродинамических процессов, определяющих успех афферентного синтеза, является нарушенным.
Но мы знаем, что может оказаться ненарушенным ни один из компонентов афферентного синтеза и больной принимает вполне адекватное решение и выполняет соответствующее движение. Однако у этих больных может быть нарушен очень важный завершающий механизм системы: оценка результатов действия в акцепторе результатов действия. В этом случае мы видим, что, правильно выполняя инструкцию, больной не может остановиться на правильно выполненном результате и продолжает его выполнять повторно.
Физиологически это значит, что больной не может оборвать действие при правильно полученном результате только потому, что аппарат сличения и, следовательно, торможения дальнейшего повторения оказывается неполноценным (систематические исследования А. Р. Лурия).
Можно назвать многие другие области научного знания, которые получили с применением теории функциональной системы иное и более конструктивное решение, чем при обычном подходе. Так, например, в корне изменилась расшифровка компенсаторного процесса при восстановлении нарушенных функций, более глубоко понят генез гипертензивных состояний вообще и, в частности, гипертонической болезни, изменилась расшифровка понятий заболевания и выздоровления и т. д.
Практически все формы нарушений и нормальные функции организма значительно более легко расшифровываются с тонки зрения теории функциональной системы, чем при обычном подходе. И здесь, конечно, на первом плане стоит расшифровка самого поведения животного и человека.
Каждый поведенческий акт, приносящий какой-то результат, большой или малый, неизбежно формируется по принципу функциональной системы.
И, наконец, решительно изменилось понимание онтогенетического развития функций и механизмов отбора на протяжении филогенеза животных.
Приведенные выше примеры являются далеко не единственными, и заметный практический успех во всех этих случаях делает особенно необходимой глубокую и всестороннюю разработку теоретических основ функциональной системы.
Обратимся к вопросу о развитии системного подхода в Советском Союзе. Нам кажется, что попытки популяризировать системный подход как новую форму трактовки и вообще подхода к научным фактам весьма полезны. Мы должны воооружить нашу научно-исследовательскую молодежь этим методом и дать ей возможность работать в ускоренном темпе, с более рациональным исходом.
Однако надо отметить, что публикации последнего года, относящиеся к этой проблеме, весьма мало сдвигают проблему системного подхода именно в сторону конструктивных методов исследования. Виной этому, как мы уже отмечали, излишнее теоретизирование конкретных биологических, машинных или общественных систем.
Общая теория систем может быть интересна исследователям в области конкретных наук только в одном плане, именно в обогащении приемов и подходов к конкретному научному исследованию. В познавательном же отношении максимальный интерес и максимальное внимание должны заслужить те концепции, гипотезы и теории, которые показали совершенно очевидный конструктивный эффект в конкретном научном исследовании.
Можно только вкратце отметить важнейшие обобщения, которые нами были разработаны с помощью общей теории функциональных систем, обобщения, которые не могли быть получены иначе. Прежде всего это концепция системогенеза, которая сейчас помогла понять основной закон эмбрионального созревания функций, обеспечивающих выживание новорожденного (П.
К. Анохин, 1937, 1948, 1964, 1968).
Концепция системогенеза по своей сути вступает в противоречие с общепринятым понятием органогенеза и дает реальное объяснение многому, что не находило до этого достаточно четкой расшифровки. Многочисленные работы наших сотрудников, начиная с 1932 г., охватывают эмбриогенез животных самых разнообразных видов, включая и изучение живых плодов (рыб, амфибий, птиц, млекопитающих, человека) (Я.
А. Милягин, К. В. Шулейкина, Е. Л. Голубева и др.).
Особенно интересно подчеркнуть, что приложение новой концепции системогенеза к онтогенетическому развитию животных разных видов дало возможность по-новому подойти к некоторым нерешенным проблемам теории естественного отбора и биогенетического закона Мюллера-Геккеля (Ф. А. Ата-Мурадова, 1970).
Эта широкая перспектива приложения системогенеза к раскрытию некоторых сторон фундаментальных проблем биологии сделала эту концепцию одним из отправных пунктов для построения теоретической биологии (П. К. Анохин, 1970).
стадии созревания плода, когда отдельные фрагменты функциональной системы (мышцы, нервные центры, кости и др.) закладываются раздельно и развиваются, так сказать, навстречу друг другу до момента их функциональной консолидации. Последняя обычно происходит за несколько дней или недель до рождения, и к моменту рождения система готова функционировать и обеспечить новорожденному выживание.
Рано или поздно должен возникнуть вопрос о том, какие факторы в раннем эмбриогенезе направляют возникновение и развитие так гармонически сочетающихся впоследствии компонентов системы. Этот вопрос подвел нас вплотную к закономерностям общей генетики, ибо только она может помочь решить кардинальный вопрос развития: как закодированы функциональные системы в гепомных организациях и какой фактор в дальнейшем дирижирует таким синхронным объединением всех компонентов функциональной системы (гетерохрония в закладках компонентов и синхронность в консолидации к моменту рождения).
Для решения этих вопросов в настоящее время мы вступили в рабочий контакт с Институтом общей генетики АН СССР и надеемся, что новый вопрос о генетике конкретных функциональных систем организма будет в какой-то степени освещен.
Другим значительным обобщением, вытекающим из физиологической архитектоники функциональной системы, является формулировка интегративной деятельности нейрона, построенная на совершенно иных основаниях, чем это принято в современной нейрофизиологии.
Эта новая концепция возникла как следствие тонкого анализа механизмов афферентного синтеза, являющегося аванпостным узловым механизмом в развертывании функциональной системы.
В самом деле, суть афферентного синтеза состоит в том, что процессы возбуждения различного функционального смысла и различного пространственного рецепторного происхождения должны быть неминуемо обработаны совместно и часто одновременно.
Возникает критический вопрос, где может быть организована эта встреча возбуждений, составляющих афферентный синтез функциональной системы.
Ответ может быть только один: такая встреча может произойти на одном и том же нейроне, сколько бы ни было переключений и обогащений этих возбуждений на пути к коре головного мозга. Отсюда возникла целая серия работ, которая привела вначале к формулировке идеи о гетерохимическом характере субсинаптических образований и новой концепции представление об интегративной деятельности нейрона (П.
К. Анохин, 1974).
Сейчас можно лишь перечислить те новые направления, которые возникли у нас в лаборатории и которые, как правило, повели мысль по новому пути.
Совсем другой аспект возник в объяснении природы вызванного потенциала, и, что особенно важно, он перестал быть только электрическим феноменом с изменяемыми параметрами: амплитудой, частотой, длительностью латентного периода и др. Благодаря системному подходу вызванный потенциал стал инструментом нейрофизиологического анализа субкортикальных взаимодействий, а вместе с тем дал возможность сделать оценку восходящих возбуждений этих подкорковых образований (Ф.
А. Ата-Мурадова).
Можно назвать области физиологии, которые получили от общей теории
функциональных систем новое направление и в объяснении и в разработке. Сюда относятся компенсация нарушенных функций, гипертоническая болезнь, эмоциональные стрессы и т. д.
Если к этому прибавить использование теории функциональных систем педагогами, медиками, музыкантами и многими другими специалистами, то можно достаточно уверенно утверждать, что в общей теории функциональных систем были найдены универсальные черты функционирования, изоморфные для огромного количества объектов, относящихся к различным классам явлений.
Это обстоятельство полностью удовлетворяет тем требованиям, которые мы предъявили в самом начале этой статьи к теории систем вообще и общей теории функциональных систем в частности.
ЛИТЕРАТУРА
Акоф Р. Л. Системы, организации и междисциплинарные исследования. В кн.: Исследования по общей теории систем.
М., 1969, с. 143144.
Анохин П. К. Проблема центра и периферии в современной физиологии нервной деятельности. В кн.: Проблема центра и периферии в нервной деятельности.
Горький, 1935, с. 970.
Анохин П. К. Системогенез как общая закономерность эволюционного процесса. Бюлл. экспер. биол., 1948, т. 26, вып.
2, 8, с. 8199.
Анохин П. К. О физиологическом субстрате сигнальных реакций. Журн. высш.
Нерв. деят., 1957, 7, 1, с. 3948.
Анохин П. К. Методологическое значение кибернетических закономерностей. В кн.: Материалистическая диалектика и методы естественных наук.
М., 1968, с. 547 587.
Анохин П. К. Функциональная система, как методологический принцип биологического и физиологического наследования. В кн.: Системная организация физиологических функций.
М., 1968, с. 57.
Анохин П. К. Теория функциональной системы. Успехи физиол. наук, 1970, т. 1, 1, с. 19-54.
Анохин П. К. Системный анализ интегративной деятельности нейрона. Успехи физиол. наук, 1974, т. 5, 2, с. 592,.
Анохин П. К., Стреж Е. Изучение динамики высшей нервной деятельности. 6. Характеристика рецептивной функции коры больших полушарий в момент действия безусловного раздражителя.
Физиол. журн. СССР, 1934, т. 17, 5, с. 1225 1237.
Ата-Мурадова Ф. А. Анализ вызванных ответов коры мозга с помощью локального теплового блока ее поверхностных слоев. Физиол. журн.
СССР, 1966, т. 52, 6, с. 696708.
Ата-Мурадова Ф. А. Гетерохронное созревание отрицательных компонентов вызванного потенциала зрительной воры в онтогенезе кролика. В кн.: Электрофизиологические исследования центральной нервной системы позвоночных.
Л., 1970, .85-92.
Афанасьев В. Г. Управление обществом как социологическая проблема. В кн.: Научное управление обществом. Вып.
2. М., 1968, с. 183224.
Гвишиани Д. М., Микулинский С. Р., Ярошевский М. Г. Социальные и психологические аспекты изучения деятельности ученого. Вопр. философии, 1971, 3, с. 83-92.
Глушков В. М, Информация по кибернетике (Институт кибернетики Украинск. GCP).
Киев, 1970.
Гудвин Б. Временная организация клетки. М., Мир, 1966, 251 с.
Дубинин Н. П. Общая генетика. М., Наука, 1970, 487 с.
Журавлев В. В. Особенности реакций нейронов гипоталамуса у голодных кроликов на пиЩ'ввое подкрепление. Тр.
Ин-та норм, и пат. физиол. АМН СССР, 1969, т. 12, с. 194196.
Зачиняева И. А. Выработка динамического стереотипа на двигательных условных реакциях. В кн.: Проблемы высшей нервной деятельности. М., Изд-во АМН.
1949, с. 521539.
Кузнецов Б. Г. Этюды об Эйнштейне. М., Наука, 1970.
Кухтин В. С. Системно-структурный подход и специфика философского знания. Вопр. философии, 1968, 11, с. 4758.
Лобные доли и регуляция психических процессов. Нейропсихологнческие исследования. М., Изд-во МГУ, 1966, 740 с.
Макаров В. А. Роль миндалевидного комплекса в механизме конвергенции возбуждений различной сенсорной модальности на нейронах коры больших полушарий. Докл.
АН СССР, 1970, т. 194, 6, с. 14541457.
Месарович М. Д. Теория систем и биология: точка зрения теоретика. В кн.: Системные исследования.
М., 1970, с. 137163.
Павлов И. П. Рефлекс цели. В кн.: И. П. Павлов. Полное собрание трудов.
Т. 3. М.-Л., 1949, с. 242347.
Павлов И. П. Динамическая стереотипия высшего отдела головного мозга. В кн.: И. П. Павлов.
Полное собрание трудов. Т. 3. М.Л., 1949, с. 496499.
Полянцев В. А. Методика самоуправляемого искусственного дыхания. Пат. физиол. и экспер. терапия, 1961, т. 5, N 4, с. 7779.
Правдивцев В. А. Эфферентно-афферентная конвергенция на нейронах зрительной коры. В кн.: Интегративная деятельность мозга.
М., 1967, с. 6769.
Раппопорт А. Математические аспекты абстрактного анализа систем. М., Прогресс, 1969, с. 83 105.
Садовский В. Н., Юдин Э. Г. Задачи, методы и приложения общей теории систем (Вступ. статья). В кн.: Исследования по общей теории юистем.
М., 1969, с. 322.
Синичкин В. В. Особенности распространения возбуждения, вызванного антидромным раздражением пирамидного тракта на нейроны зрительной области коры головного мозга. В кн.: Интегративная деятельность мозга.
М., 1967, с. 7274.
Системные исследования. Ежегодник (Институт истории естествознания и техники АН СССР). М., Наука, 1970, 208 с.
Судаков К. В. Нейрофизиологические механизмы пищевого возбуждения. Докт. дисс. М., 1965, 488 к.
Судаков К. В. Пейсмекерный механизм пищевой мотивации. Рефераты докл.
11-го Всесоюзн. съезда физиологов. Т. 1. Л., 1970, с. 187492.
Судаков К. В., Котов А. В. Взаимодействие мотивационного и подкрепляющего возбуждений на нейронах коры мозга как основа пищевого условного рефлекса. Материалы симпозиумов 22-го совещания по проблемам физиол. высшей нервной деятельности.
Л., .1969, с. 2021.
Судаков К. В., Фадеев Ю. А. Особенности восходящей активности коры головного мозга в состоянии физиологического голода и при болевом раздражении. Физиол. журн.
СССР, 1963, т. 49, И, с. 13101317.
Трапезников В. А. Человек в системе управления. Тезисы 5-го Всесоюзн. совещания по проблемам управления.
М., 1971.
Урсул А. Д. Природа информации. М., Политиздат, 1968, 288 с.
Ухтомский А. А. Собр. соч. Т. 2. Л., Изд-во АН СССР, 1951, 180 с.
Хаютин С. Н. Активность одиночных нейронов зрительной области коры в условиях (выраженной пищевой мотивации. В кн.: Структурная, функциональная и нейрохимическая организация эмоций.
Л., 1971, с. 175,179.
Шумилина А. И. Функциональное значение лобных областей коры головного мозга в условнорефлекторной деятельности собаки. В кн.: Проблемы высшей нервной деятельности.
М., 1949, с. 561627.
Шумилина А. И. Экспериментальный анализ методом вызванных потенциалов корково-подкорковой реверберации возбуждений. В кн.: Интегративная деятельность нервной системы в норме и патологии.
М., 1968, с. 276287.
Эйнштейн А. Физика и реальность. М., Наука, 1965, 359 с.
Функциональная система как основа интеграции нервных процессов в эмбриогенезе. Сборник докл.
6-го Всесоюзного съезда физиологов, биохимиков и фармакологов. Тбилиси, 1937, с. 148 156.
Авт.: П. К. Анохин, А. И. Шумилина, Я. А. Милягин, Е. Л. Гонубева и др.
(Ashby W. R.) Эшби У. Р. Конструкция мозга. Пер. с англ. М., Мир, 1964, 411 с.
(Aschby W. R.) Эшби У. Р. Общая теория систем как новая научная дисциплина. В кн.: Исследования по общей теории систем.
М., ,1969, с. 125,142.
(Bertalanffy L.) Берталанфи Л. Общая теория систем критический обзор. В кн.: Исследования по общей теории систем.
М., 1969, с. 2324.
Brass J. D. Design for decision. New York, 1953.
Bullock T. H. Parameters of integrative action of the nervous system at the neuronal level. In: The submicroscopic organization and function of nervous cells 1958, p. 323.
Калман Р., Фалб П., Арбиб М. Очерки по математической теории систем. Пер. с англ. М., Мир, 1971, 400 с.
Ханике Ф. Новые идеи в области управления. Пер. с англ.
М., Прогресс, 1969, 125с.
Полл А. Д., Фейджин Р. Е. Определение понятий Системы. В кн.: Исследования общей теории систем.
М., 1969, с. 252282.
Юнг Р. Ярче тысячи солнц. Пер. с англ. М., Атомиздат, 1960, 280 с. (Lange О.) Ланге О. Целое и развитие в свете кибернетики.
В кн.: Исследования по общей теории систем. М., 1969, с. 18І251.
Marling D. С. La experimentation instrumental a la luz de conception de Anojin.
Bol. Inst. neurofisiol. psycol., 1966, v. 2, p. 14. Милсум Дж. Анализ биологических систем управления.
Пер. с англ. М., Мир, 1968, 501 с.
Паек Г. Естественная теория цепей. В кн.: Саморегулирующая система. М., 1964, с. 318357.
Suga N. Recovery cycles and responses to frequency modulated tone pulses in auditory neurones of echo-locating bats. J. PhysioL, 1964, v. 175, p. 5080.
(Waddington С. Н.) Уоддингтон К. В кн.: Б. Гудвин. Временная организация клетки. М., 1966.
Предисловие.
(Weiss Р.) Вепсе П. Динамика клетки. В кн.: Современные проблемы биофизики.
Т. 1. М., 1961, с. 2134.